
林幸助
國立中興大學生命科學系終身特聘教授、環境保育暨防災科技研究中心主任,國立台灣海洋大學海洋環境與生態研究所合聘教授,國立中山大學海洋科學系合聘教授,國際自然保護聯盟(IUCN)紅樹林生態系紅皮書領銜作者、紅樹林專家群專家,聯合國環境規劃署(UNEP)生物多樣性和生態系統服務政府間科學與政策平台(IPBES)亞太地區生物多樣性和生態系統服務評估報告領銜作者。專長海草床、珊瑚礁、紅樹林、河口、潟湖、湖泊、濕地與溪流生態系統結構與功能與服務。2022年榮獲教育部海洋教育推手獎及國際永續碳中和研究獎 (The Sustainability 2022 Carbon Neutrality Award),2023年以「自然碳匯量測技術開發與推動策略」榮獲國家農業科學獎。
第 28 屆聯合國氣候大會(COP28)於 2023 年 12 月杜拜落幕,會員國經過長時間的談判終於達成「阿聯共識」(UAE consensus),同意「以公正、有序且公平」的方式「脫離」化石燃料,以符合科學的方式在 2050 年達成淨零排放目標。在為期兩週的會議期間也產生許多協議,其中之一是 12 月 10 日由 40 個國家或地方實體簽署支持阿聯所發起的「紅樹林突破」(Mangrove Breakthrough)專案,目標期望在 2030 年前投注 40 億美元,復育和保護全球 1,500 萬公頃的紅樹林,遏止紅樹林的毀林行為。
碳吸存速率優於森林!紅樹林生態系服務價值高
紅樹林生長於熱帶與亞熱帶地區,提供豐富的生態系服務,與海草床、鹽沼並列為三大濱海「藍碳」生態系。國際自然保育聯盟(IUCN) 2020 年估計紅樹林生態系服務價值高達每年 8,000 億美元,這些服務包括穩固海岸、防止侵蝕、淨化水質、防風與減少暴潮海浪的衝擊,是海岸的天然屏障;紅樹林也具有高生產力及豐富生物多樣性,提供豐富的食物與複雜的物理結構,為海洋動物提供良好的生長與養育場域;紅樹林木材本身能作為木炭燃燒或建築使用,木材碎片則能製成紙漿的原料,樹皮可提取單寧酸作為醫藥與鞣製皮革和染料之用;紅樹林也能為人類提供文化美學與休憩娛樂服務等。
紅樹林碳匯(Carbon sink)功能在近 10 年備受關注,因為紅樹林的「藍碳」具有比稱為「綠碳」的陸域森林高 2~3 倍的碳吸存速率,主要原因是紅樹林生長的沿岸河口環境營養豐富,競爭者少。「綠碳」與「藍碳」的碳儲存量與方式有很大差異。一般森林儲存在植物體或土壤的碳儲存量大致相等,但兩者加總起來的總碳儲存量只有紅樹林碳儲存量(每公頃高達 1,500 公噸)的四分之一。「藍碳」比「綠碳」能更有效率儲存有機碳的原因,除了因為光合作用速率較快之外,另一個原因是「藍碳」發生森林野火的風險低。森林野火發生將使得原本儲存在樹幹及土壤的有機碳會被釋放回大氣中。紅樹林的高碳吸存速率,大量的枯枝落葉及死亡根系經分解後會累積於土壤中,因土壤長期保持厭氧條件,也可減緩土壤有機碳的分解速率,因此碳儲存在紅樹林土壤的比例很高,大約是 80%,而儲存在植物體的比例只有 20%。
2022 年12 月 7日至19 日於加拿大蒙特婁舉行之聯合國第 15 屆生物多樣性大會(CBD COP 15),「以自然為本解決方案」(Nature-based Solutions,NbS)成為大會焦點,也是未來 10 年全球保育主流。NbS 指的是可有效調適與回應社會挑戰,作為永續管理及復育自然生態系之保護行動,可同時提供人類福祉和生物多樣性效益。因此 CBD COP 15 呼籲各界運用自然方式來減碳或吸碳,以調適氣候變遷的影響。世界自然基金會(World Wild Fund,WWF)指出,高達 92% 國家氣候承諾(或稱為國家自主貢獻,NDCs)都納入自然解方,期望運用自然碳匯方式來減排並調適氣候變遷的影響。紅樹林碳匯即是以自然為本的解決方案,成本低但效益高,因此「自然碳匯」也是台灣 2050 淨零轉型十二項關鍵戰略的第九項。2021 年 6 月,聯合國政府間生物多樣性與生態系服務科學政策平台(IPBES)和政府間氣候變化專門委員會(IPCC)共同指出,紅樹林本身是碳匯,同時也是海岸生物多樣性的熱點,因此保護紅樹林,等於同時保護生物多樣性與碳匯是雙贏策略。「紅樹林突破」專案將有助於將資金導入紅樹林復育與保育,這對於實現第 21 屆聯合國氣候大會(UNFCCC COP21)之《巴黎協定》控制全球升溫 1.5℃和第 15 屆生物多樣性大會(CBD COP 15)之《昆蒙全球生物多樣性框架》目標至關重要。

台灣紅樹林持續增加,北廻歸線以南多樣性高
紅樹林為一水生木本植物之類群統稱,全球紅樹林樹種共有 17 科 30 屬 81 種,面積共 14 萬平方公里(即1,400萬公頃),主要以紅樹科植物為代表,一般須具有胎生及呼吸根的構造,並且能夠適應半淡鹹水交會之環境。紅樹林雖然只占全球熱帶森林面積的 0.7%,但是毀損紅樹林後所導致的碳排放量相當於全球砍伐森林碳排放量的 10%。
在台灣的紅樹林樹種紀錄有 6 種,包括馬鞭草科的海茄苳(Avicennia marina)、紅樹科的五梨跤(Rhizophora stylosa)、水筆仔(Kandelia obovata)、使君子科的欖李(Lumnitzera racemosa)、紅茄苳(Bruguiera gymnorrhiza)及細蕊紅樹(Ceriops tagal)。其中紅茄苳與細蕊紅樹已自自然棲地滅絕。台灣紅樹林的分布樹種在北迴歸線以北趨向單一,以南組成較為多元。水筆仔與海茄苳為台灣紅樹林中較優勢的樹種,桃園以北的紅樹林均屬水筆仔純林,而彰化芳苑則為水筆仔成林的最南界,於台灣熱帶地區數量極少。新竹新豐為台灣海茄苳分布的最北界,而南至屏東大鵬灣均有分布。欖李及五梨跤兩樹種成林現象相較另外兩種少見,僅出現於嘉義以南的地區。欖李普遍分布於高程較高的區域,五梨跤普遍分布於高程較低的區域,目前以好美寮、雙春的五梨跤族群最大。
1950 年文獻紀載,台灣紅樹林分布於基隆港、台北淡水、新竹新豐、台南、高雄左營、高雄港、屏東東港及車城 8 區。其中,基隆及車城為當時紅樹林分布的最北及最南界,而高雄港紅樹林物種多樣性最高,亦是當時唯一 6 種紅樹林(包括已滅絕紅茄苳與細蕊紅樹)均有紀錄的分布地,而左營亦有紅茄苳之紀錄。基隆港及高雄港的紅樹林因港口開發而消失,也導致原生的紅茄苳與細蕊紅樹就此在台灣消失。台灣紅樹林於 1976~2011 年間在台灣本島的面積由 178 公頃增加至 586 公頃,而於 2019 年調查面積共 660.7 公頃,約增加 271%,加上離島地區的調查,總面積達到 680.7 公頃。紅樹林面積隨年代持續增加,於 1996 年後有顯著增加,應與國內保育意識抬頭與行動崛起有關,但在 2011 年之後增加的趨勢開始減緩,此減緩的現象應與大規模的紅樹林移除計畫啟動有關。

目前紅樹林面積最大者為台南市,面積為 226.2 公頃,占總面積約 33.9%,面積增加較為明顯主要為沿急水溪的北門至學甲的紅樹林擴張。其次為淡水河流域涵蓋的新北市與台北市,總面積為 125.6 公頃,占總面積約 18.8%,其中蘆洲為近年新擴張之區域,目前面積約 20.2 公頃。嘉義在 1986~1996年區間紅樹林面積減少,主要因為台鹽公司於好美寮開展機械化鹽灘,在紅樹林中間建造土堤,造成紅樹林大量死亡,後續此區紅樹林因復育而逐漸恢復,再加上朴子溪的紅樹林持續擴張,使嘉義市之紅樹林面積逐年增加。新竹紅樹林面積雖逐年上升,但在 2019 年面積大幅下滑,主因為香山地區的紅樹林大規模移除行動。彰化王功的紅樹林成林亦是 5 年內擴張的結果,而高雄中都人工濕地公園之紅樹林則為自 2011 年人工種植的成果。
碳匯能力強!環境因素比樹種更決定紅樹林碳循環
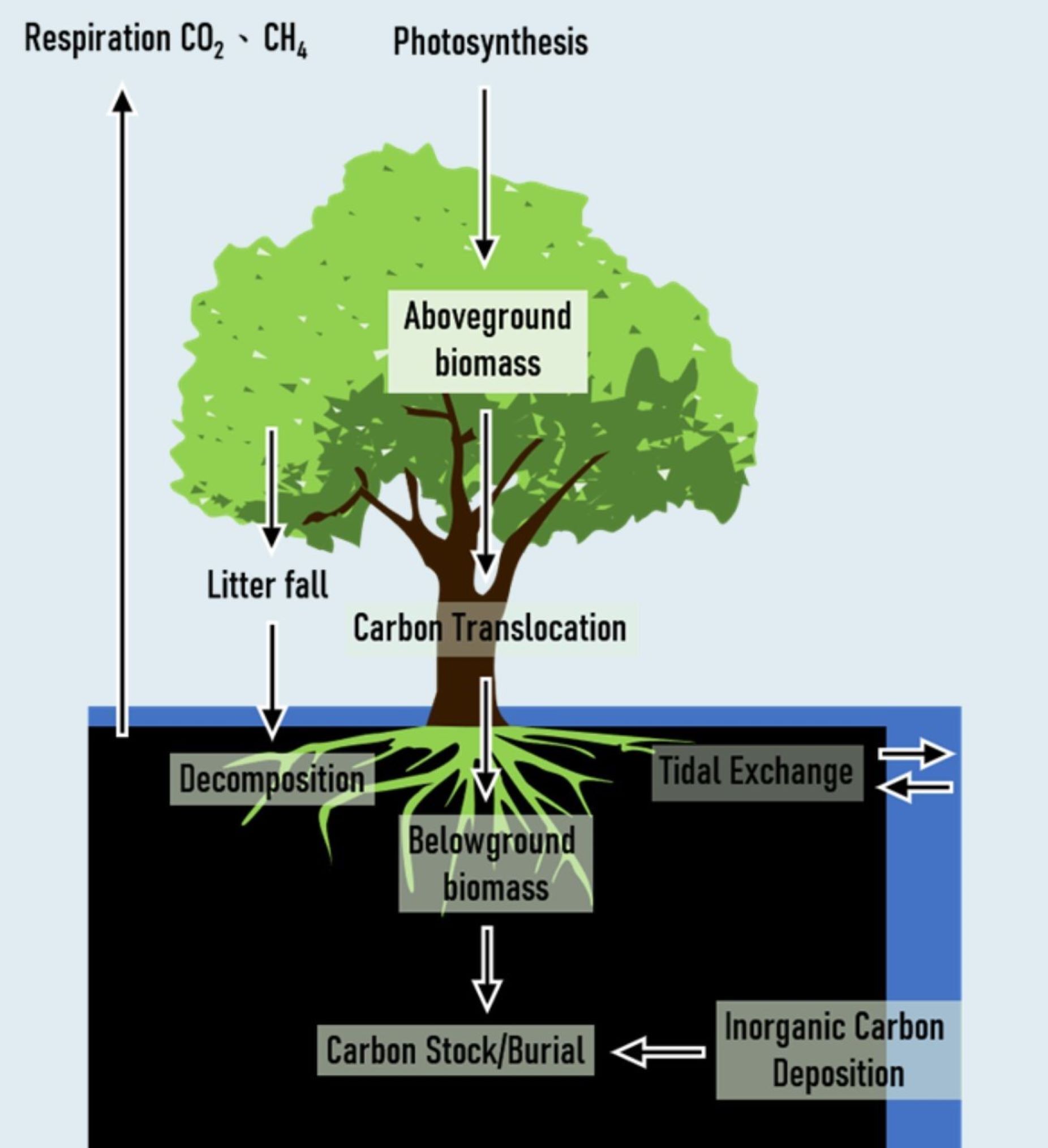
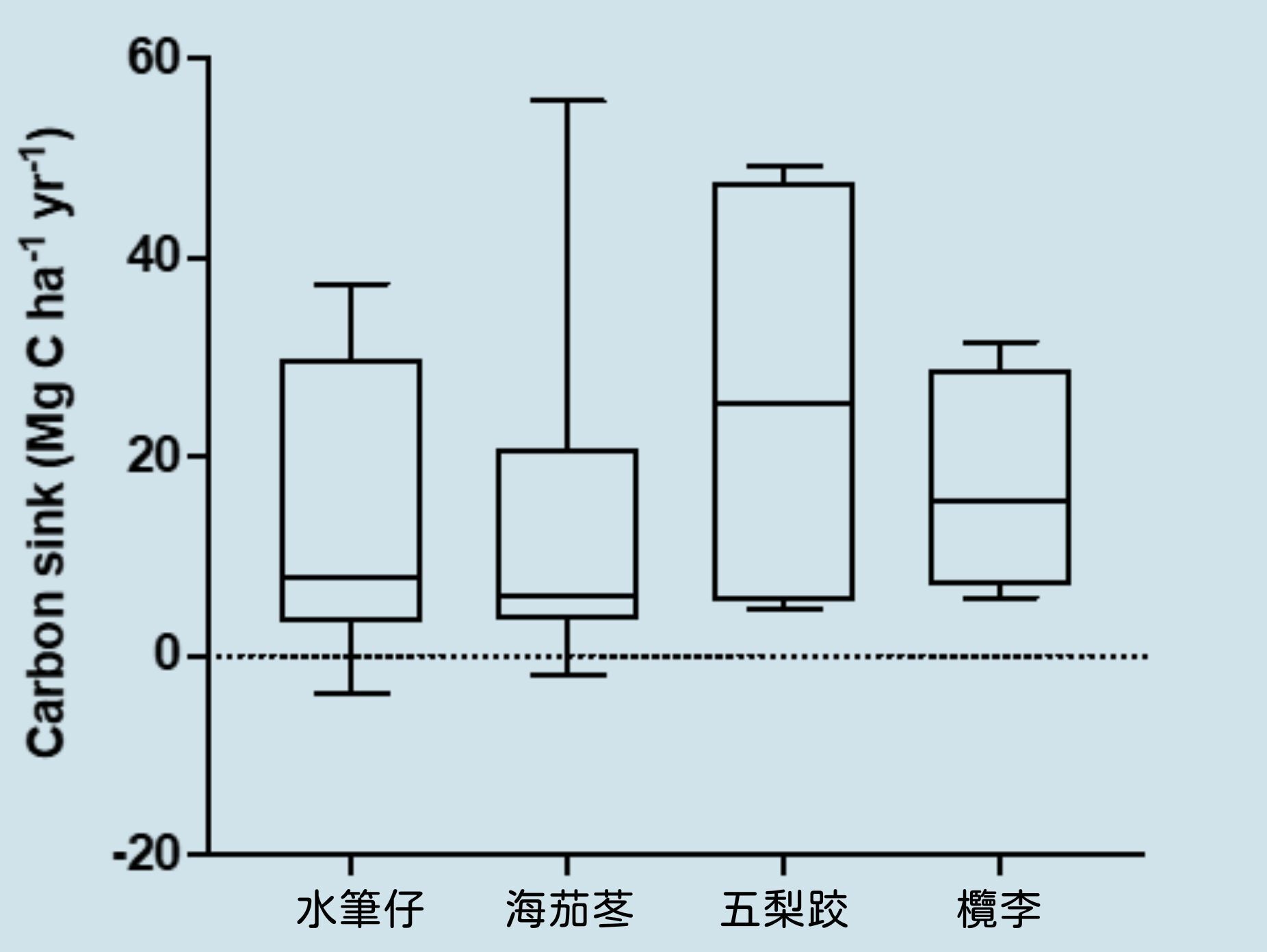
建構碳收支模式才能了解紅樹林形成碳匯(Carbon sink)的機制,也才能了解影響紅樹林儲碳量的重要因素。紅樹林碳收支模式顯示紅樹林生長行光合作用(Photosynthesis)將大氣中 CO₂ 固定於植物體內,除了產生地上部生物量(Aboveground biomass),透過碳移轉(Carbon translocation)也會生產地下部生物量(Belowground biomass),以及死有機碳枯落物(Litter fall)。這些有機碳扣除微生物分解作用後才能埋藏儲存於土壤中(Carbon burial)形成儲碳量(Carbon stock)。來自於其他棲地的有機碳或無機碳(inorganic carbon)也會藉由潮汐海流之橫向交換(Tidal exchange)沉積於紅樹林底土中。底土中異營微生物則會進行呼吸作用將 CO₂、甲烷(CH₄)及氧化亞氮(N₂O)等溫室氣體釋放回空氣中。台灣水筆仔、海茄苳、五梨跤及欖李等四種紅樹林的碳吸存速率估計分別為 13.73、13.36、26.23 及 19.23 Mg C ha⁻¹ yr⁻¹ [1],以全台分布面積(680.7 ha)計算紅樹林碳匯量為 17,497 Mg C yr⁻¹ ,顯示台灣紅樹林的碳匯能力不容小覷,應該及早納入國家溫室氣體排放清冊之國家自主貢獻(NDC)內。
紅樹林常受到人類活動所排放的淡水或營養污水影響,而改變其碳吸存速率,尤其以氮源輸入對於紅樹林碳匯能力影響最劇。其中最受注目的是溫室氣體 CH₄ 及 N₂O,因為同樣重量,CH₄ 的暖化潛勢是 CO₂ 的 28 倍,而 N₂O 的暖化潛勢是 CO₂ 的 298 倍之多。由於微生物進行氮循環反應的各階段所能獲得的能量皆不同,因此微生物在富營養的污水環境裡,常會造成不完全的硝化或脫氮反應,使 N₂O 以中間產物的形式直接排入大氣中,可能會因此削減紅樹林碳儲存效益。
紅樹林也可能受到人為活動 (如土地利用或地景改變、河川或海岸工程及污染等)及氣候變遷(如海平面上升、暖化、酸化及極端氣候等)的影響。全球 1980~2000年間消失的紅樹林面積高達 35%,每年消失的紅樹林超過 28 萬公頃,其中人為活動的直接與間接影響,是導致全球紅樹林面積急遽減少的主因。紅樹林也是受到海平面上升等氣候變遷影響最劇烈的生態系統之一。因其位於河口潮間帶或海岸地區,因此近年來常出現海岸藍碳系統的地形抬升速度能否趕上海平面上升速度的脆弱度研究。絕大部分的紅樹林復育計畫由於忽略了水文與紅樹林間的關係,不是以完全失敗收場,就是未能達到當初設定的目標。紅樹林因具備複雜的根系、枝幹、樹冠層結構,故可顯著影響水流特徵及泥砂有機質沉積過程,進而影響地形地貌之變化速度。這些複雜的交互作用可能影響紅樹林內部的碳儲存收支情形。而河口及海岸帶的紅樹林有助於穩固由流域上游排放下來的泥砂,可能有助於減緩海岸地區對抗海平面上升的威脅。
台灣紅樹林應多元共治,創造多樣化地景
1995 年至今,台灣紅樹林雖少有因大型開發案而消失之現象,但不同地區的紅樹林卻因海岸河口之變動而有不同的發展。如在淡水河口之紅樹林逐漸擴散與陸化,影響區域排水與生物多樣性,而新竹香山濕地則因海茄苳紅樹林蔓生影響沿海濕地生態。新竹市政府於 2015 年底以大型機具進入沿海濕地砍除紅樹林海茄苳,並採就地掩埋方式清除紅樹林,困擾居民的小黑蚊已減少,看起來濕地原有的蟹類與水鳥數量逐漸增加,但有些依附紅樹林生長的生物卻有可能消失,碳吸存能力也會大幅下降。但在濁水溪以南沿海地區則因地層下陷、突堤效應及灘地流失等因素,大面積紅樹林消失,顯示台灣紅樹林生態系南北面臨不同的危機。
因此,除了需要因地制宜推動紅樹林伐除或復育計畫,更要從宏觀角度思考紅樹林伐除或復育對整體生態系服務的影響,不宜僅以單一物種之保育為考量。應該將各沿海濕地自然資源的治理層級,推向社區與國家共同管理的思考層級。面對過度生長之紅樹林若能有一定程度的移除,可能可以創造更多樣化的地景,提昇生物多樣性,但須要儘量減少碳匯功能的減損,維護整體生態系功能與服務;而在原有紅樹林逐漸單一化或消失的棲地,則應考量復育多樣化之紅樹林樹種或棲地,以有效復育生物多樣性與生態功能,提昇生態系服務與碳匯能力。但無論是進行復育或伐除的管理,都必須考量公共資源旳社會應對面,才能實踐當前國際倡議紅樹林管理思維下的多元共治理念。
刊出時間:2024 年 1 月 23 日
關鍵字:紅樹林、藍碳、氣候變遷、NbS、生物多樣性